
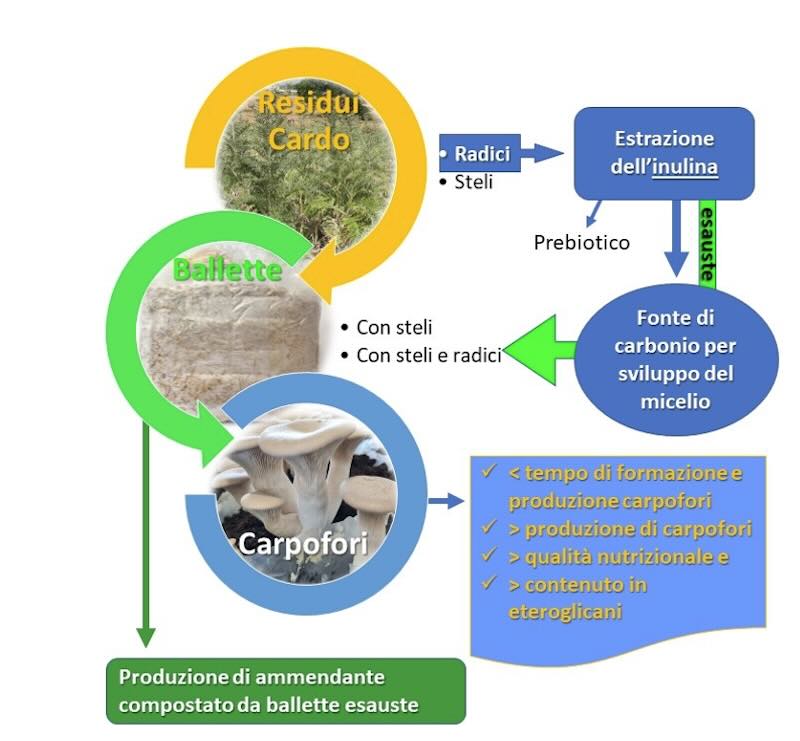
I funghi sono alimento consumato in tutto il mondo e sono apprezzati, per le loro qualità organolettiche, salutari per la dieta umana e come fonte di proteine. I metaboliti ottenuti dai miceli e dai corpi fruttiferi di alcune specie fungine hanno molteplici attività farmacologiche e molti sono disponibili in commercio come integratori. La coltivazione in Italia del cardo può rappresentare un'opportunità data la sua versatilità, potendo ottenere dalla coltivazione diversi bioprodotti (bioplastica, biopesticidi, biocosmetici, bioenergia, eccetera) e utilizzandone poi la biomassa residua, rappresentata da fusti e radici con alto contenuto di inulina oltre che biotina e tiamina due vitamine spesso aggiunte ai substrati, per la coltivazione di funghi eduli. In Italia sono impiegati in agricoltura due dei prodotti derivati dal cardo la bioplastica per la pacciamatura biodegradabile e l'acido pelargonico utilizzato come diserbante, spollonante.
I basidiomiceti appartenenti al genere Pleurotus hanno un elevato contenuto proteico e sono caratterizzati da un profilo di fibra alimentare unico (principalmente β-glucano ramificato), alti livelli di vitamine del gruppo B, vitamina D, Fe, Zn, Cu, Se e alcuni composti bioattivi. Inoltre, sono anche noti per le proprietà medicinali quali attività antitumorali, antimicrobiche, antidiabetiche, anti ipercolesterolemiche e immunomodulanti. Il P. eryngii ha il più alto contenuto di glucano che lo candida come alimento funzionale e come fonte di glucano per l'industria alimentare. I glucani sono polisaccaridi, componenti strutturali delle pareti cellulari dei funghi e sono noti per la loro attività immunomodulante (Carrieri et al., 2017).
In Italia la melassa è la fonte di carbonio impiegata per la coltivazione del cardoncello, la quale è importata da altri paesi da quando non vi è più la coltivazione della barbabietola da zucchero. Inoltre, i funghi eduli possono essere coltivati su diversi substrati (farine di cereali, crusca, paglia, barbabietola da zucchero) che ne influenzano la resa e le caratteristiche chimiche e nutrizionali.
Nell'ambito delle attività di ricerca provviste nel Progetto Pon - "Colture autoctone mediterranee e loro valorizzazione con tecnologie avanzate di chimica verde" - Cometa (cod. ARS01_00606) è stata impiegata la biomassa residua del cardo (Cynara cardunculus L. var. altilis), steli e radici esauste, come substrato per la coltivazione del fungo "cardoncello" (Pleurotus eryngii). La biomassa residua del cardo, costituita degli steli e delle radici dopo la raccolta dei capolini dai quali si ottengono i semi, per la coltivazione di funghi rappresenta l'implementazione di un sistema di economia circolare: uno scarto che diventa risorsa. È noto che le radici di cardo sono ricche in inulina (>40%) e, nell'Italia meridionale, il P. eryngii selvatico è comunemente chiamato "cardoncello" ciò ha fatto ipotizzare un'elevata affinità per la pianta che contiene anche inulina nelle radici passo congruente è stato sostituire la melassa con le radici di cardo come fonte di carbonio. Inoltre, P. eryngii possiede un'elevata capacità di colonizzare la lignina (Battaglia et al., 2023).
Il cardo è proposto come coltura virtuosa per un esempio totale di economia circolare
(Fonte: Crea)
In questo lavoro è stato valutato l'utilizzo della biomassa di cardo sulla resa, qualità e contenuto in eteroglicani dei corpi fruttiferi di P. eryngii; la produzione di ammendante compostato valorizzando ballette di substrato "esauste" che rappresentano un sottoprodotto di scarto da smaltire per l'impresa a conclusione del ciclo produttivo dei funghi. Il cardo è proposto come coltura virtuosa per un esempio totale di economia circolare completa con produzione di prodotti innovativi e riutilizzo totale degli scarti di ognuno dei processi di produzione, la principale delle quali, come detto, la trasformazione di scarti in proteine nobili e prodotti funzionali.
Come è stato disegnato l'esperimento
È stato selezionato un isolato di P. eryngii con maggiore affinità alla biomassa di cardo, in quanto la variabilità intraspecifica degli isolati può essere molto elevata (Battaglia et al., 2023).
Produzione dell'inoculo
Cinquecento grammi di semi di miglio, preventivamente idratati con acqua più sali di Czapek dox (Oxoid) e sterilizzati, sono stati aggiunti a substrato sterile e inoculati con 10 plug da 1 centimetro di diametro da colture fungine di 10 giorni. I sacchetti ottenuti sono stati incubati a 22 gradi fino alla completa colonizzazione (circa 20 giorni).
Tre chilogrammi di miglio sono stati pesati e lavati con acqua di rubinetto e immersi in acqua con aggiunta di 10 grammi su litro-1 di Czapek dox (Oxoid) per 24 ore a 4 gradi.
Crescita del micelio
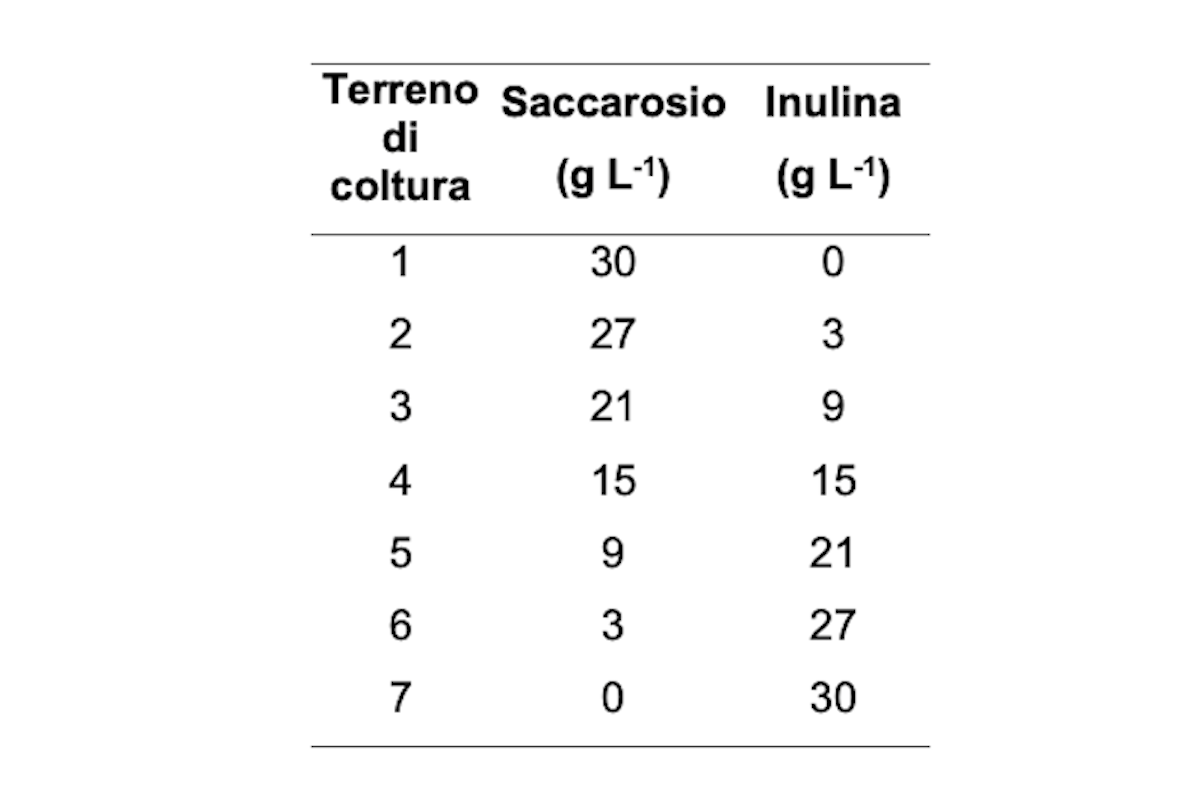
Per valutare la migliore fonte di carbonio per massimizzare la produzione del micelio di P. eryngii, sono state confrontate sette diverse concentrazioni di inulina e/o saccarosio come fonte di carbonio (Tabella 1). I terreni sono stati preparati utilizzando sali di Czapek; beute da due litri per ciascuna delle sette combinazioni di inulina/saccarosio sono state inoculate in triplicato con plug di 5 millimetri di diametro di colonie fungine di 7 giorni. Le beute sono state incubate a 22±1 gradi al buio. Il micelio è stato raccolto e pesato dopo 14 giorni; i campioni sono stati asciugati per calcolare il peso a secco (Ps). I risultati sono le medie dei tre esperimenti con quattro repliche.
Valutazione della resa e delle caratteristiche dei corpi fruttiferi su diversi substrati
(Fonte: Crea)
La produzione dei carpofori, il contenuto di eteroglicani e la composizione nutraceutica sono stati valutati mediante la coltivazione di P. eryngii su quattro diversi substrati: Cardo Steli-Inulina 600 grammi di germogli di cardo reidratati al 65-70% mediante acqua sterile ed aggiunta di 20 grammi di CaCO3 e 30 grammi di inulina; Cardo Steli-Saccarosio 600 grammi di germogli di cardo reidratati con acqua ed aggiunta di 30 grammi di saccarosio e 20 grammi di CaCO3; Cardo Steli-Radici 400 grammi di germogli di cardo stems più 200 grammi di radici di cardo reidratate con acqua sterile ed aggiunta di 20 grammi di CaCO3; Paglia frumento-Melassa 600 grammi di paglia di grano con aggiunta di 30 grammi di melassa reidratata con acqua contenente lo 0.05% NH4Cl e 20 grammi CaCO3. Tale substrato è servito come testimone.
Sono stati usati 50 grammi di miglio colonizzato con l'isolato selezionato di P. eryngii per inoculare ognuno dei 4 sacchetti e incubati a 23 gradi al buio. Dopo la colonizzazione, i sacchetti sono state aperti sulla parte superiore del sacchetto e incubati con fotoperiodo 8/16 ed è stata mantenuta un'umidità relativa fra 75 e il 90%. Sono stati valutati il tempo di colonizzazione dei sacchetti, per la differenziazione, il numero e la resa dei carpofori. Il saggio è stato ripetuto tre volte.
Determinazione degli eteroglicani nel micelio e nei carpofori
L'estrazione degli eteroglicani è stata effettuata seguendo il protocollo illustrato da Carrieri et al. (2017) determinando la concentrazione attraverso un metodo colorimetrico descritto da Nitschke et al. (2011). Sono state determinate la curva di calibrazione e la sua equazione utilizzando le molecole ottenute nel nostro studio precedente (Carrieri et al., 2017). L'assorbanza è stata calcolata come differenza tra Congo Red più eteroglicano a diverse concentrazioni e il solo colorante. Le concentrazioni riportate sono la media di due saggi con tre repliche tecniche. Il contenuto di eteroglicani ottenuto da micelio e corpi fruttiferi di P. eryngii è stato confrontato con quello ottenuto da micelio di G. lucidum allevato come riportato in Carrieri et al. (2017).
Composti bioattivi, attività antiossidante e saggi enzimatici
I composti bioattivi e l'attività antiossidante sono stati testati su estratti idroalcolici dei corpi fruttiferi prodotti su Csr e Pfm. I polifenoli ed i flavonoidi totali sono stati determinati spettrofotometricamente con il metodo Folin-Ciocalteu e del cloruro di alluminio ed i risultati calcolati come milligrammi di acido gallico equivalente (Age) per grammi di peso secco e milligrammi catechina equivalente (Ce) per grammi di peso secco. L'attività antiossidante è stata determinata con il metodo del DPPH calcolati come µmol Trolox equivalent (TE) grammi-1 di Ps.
L'attività della polifenolossidasi (Ppo) (EC.1.10.3.1) è stata misurata utilizzando una soluzione di catecolo 0.5 Moli in tampone fosfato 0.1 Moli pH 6.4 a 398 nanometri e calcolata in nanomoli su grammi-1 Ps, mentre l'attività della lipossigenasi (Lox) (EC 1.13.11.12) è stata monitorata in tampone fosfato in presenza di acido linoleico 5 microMoli a 234 nm, e determinata come in nanomoli su grammi-1 Ps. Il contenuto di proteine totali è stato determinato con il saggio Bradford.
Tutti i risultati sono espressi come incremento percentuale rispetto al campione Fpm.
Processo di compostaggio
Le ballette esauste ottenute con la biomassa residua di cardo e, dopo la produzione di carpofori di P. eryingii, ricche di micelio fungino tra cui quello di Trichoderma spp., sono state compostate in miscela con il digestato solido proveniente da un impianto di fermentazione anaerobica di effluenti bufalini. Il cumulo è stato costruito a strati alterni come si vede nella foto A, in un rapporto di 1:1 in volume che comportato la miscelazione di 100 chilogrammi di ballette esauste e 100 chilogrammi di digestato.
Il compostaggio è iniziato il 28 giugno 2022 ed è stato concluso il 10 ottobre 2022 dopo circa 100 giorni presso l'impianto sperimentale sito nel fondo agricolo annesso alla sede di Caserta del laboratorio del Crea Cerealicoltura e Colture Industriali (Crea-Ci). Il cumulo è stato insufflato di aria mediante tubazione forata posta alla sua base e collegata ad un motore soffiante, mentre la sua bagnatura è stata eseguita quotidianamente durante l'estate. Tre rivoltamenti manuali sono stati eseguiti dopo la fine della fase termofila con cadenze mensili per favorire il rimescolamento e l'ossigenazione della massa. Il 9 agosto (completata la fase termofila), il 1° settembre e l'11 ottobre, campioni di compost sono stati prelevati dal cumulo in tre punti ad una profondità di 15-20 dalla superficie, per eseguire analisi chimico-fisiche su alcuni parametri considerabili indicatori di processo: C organico, N Kjeldhal, C/N, conducibilità elettrica e pH. La temperatura nel cumulo, altro indicatore di processo, è stata misurata in continuo con una sonda collegata ad un datalogger Watch-Dog, series 1000.

Figura 1 - Preparazione del cumulo di compostaggio con il digestato (più scuro) e le ballette da funghicoltura contenenti cardo previamente frammentate
(Fonte: Crea)
Analisi dei dati
Ogni valore rappresenta la media ± errore standard (Es) di tre esperimenti indipendenti eseguiti in triplicato. Il test per l'omogeneità della varianza è stato applicato per valutare se i dati potessero essere combinati in un unico esperimento. Le analisi statistiche sono state eseguite utilizzando GraphPad InStat versione 3.00 per Windows (GraphPad Software, San Diego California Usa) e le differenze tra i trattamenti sono state considerate significative a p ≤ 0,05.
I dati ottenuti sono stati sottoposti ad analisi della varianza (Anova) per variabili quantitative e le medie sono state separate utilizzando il test di Tukey.
Risultati
Effetti sulla produzione di micelio mediante l'aggiunta di inulina e saccarosio in vitro
Sono state saggiate sette combinazioni di inulina e saccarosio al fine di verificare l'effetto dell'inulina sulla crescita del micelio e la produzione di eteroglicani nel micelio di P. eryngii. I risultati dimostrano che l'aggiunta di inulina ha influenzato positivamente lo sviluppo del micelio ottenendo un maggiore peso secco del micelio di P. eryngii (Figura 2).
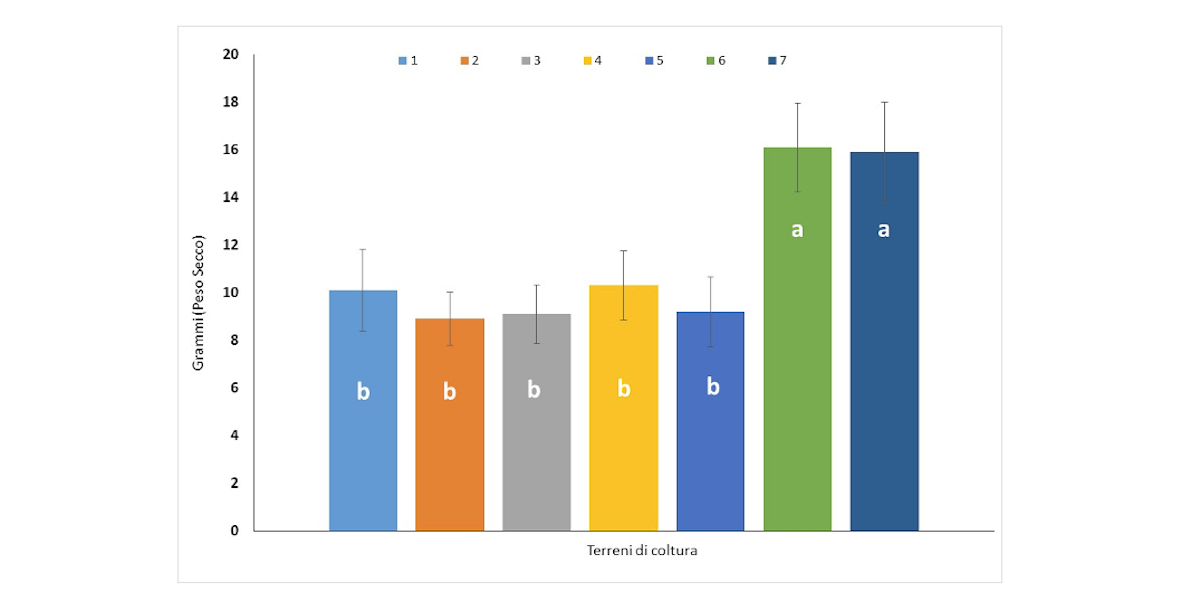
Figura 2 - Effetto dell'inulina in sette differenti terreni di coltura (cfr. tabella 1) sulla crescita del micelio di P. eryngii
Produzione dei carpofori e contenuto di eteroglicani su diversi substrati
(Fonte: Crea)
I substrati hanno influenzato la crescita del micelio che su Csr e Css ha colonizzato il panetto in 15,6 giorni, mentre per Csi e Pfm ha impiegato rispettivamente 17,4 e 19,2 giorni (Figure 3.A, 3.B). Lo sviluppo dei primordi di P. eryngii, invece, ha richiesto 22 giorni su Csi, mentre occorrono almeno 26 giorni sui substrati senza radici. Il substrato ha influenzato la velocità di formazione dei carpofori e la resa (Figura 3). La formazione dei carpofori è più veloce su Csr (5 giorni vs 7 e 8 giorni) rispetto agli altri substrati in studio.
La resa è positivamente influenzata dall'aggiunta delle radici esauste del cardo, in quanto il peso fresco medio del fungo è stato maggiore in Csr (371 g) rispetto agli altri substrati (Figure 4.A, 4.B) e, comunque il peso medio dei carpofori era sempre maggiore quando nel substrato vi era biomassa di cardo anziché la paglia di frumento (Figura 4).
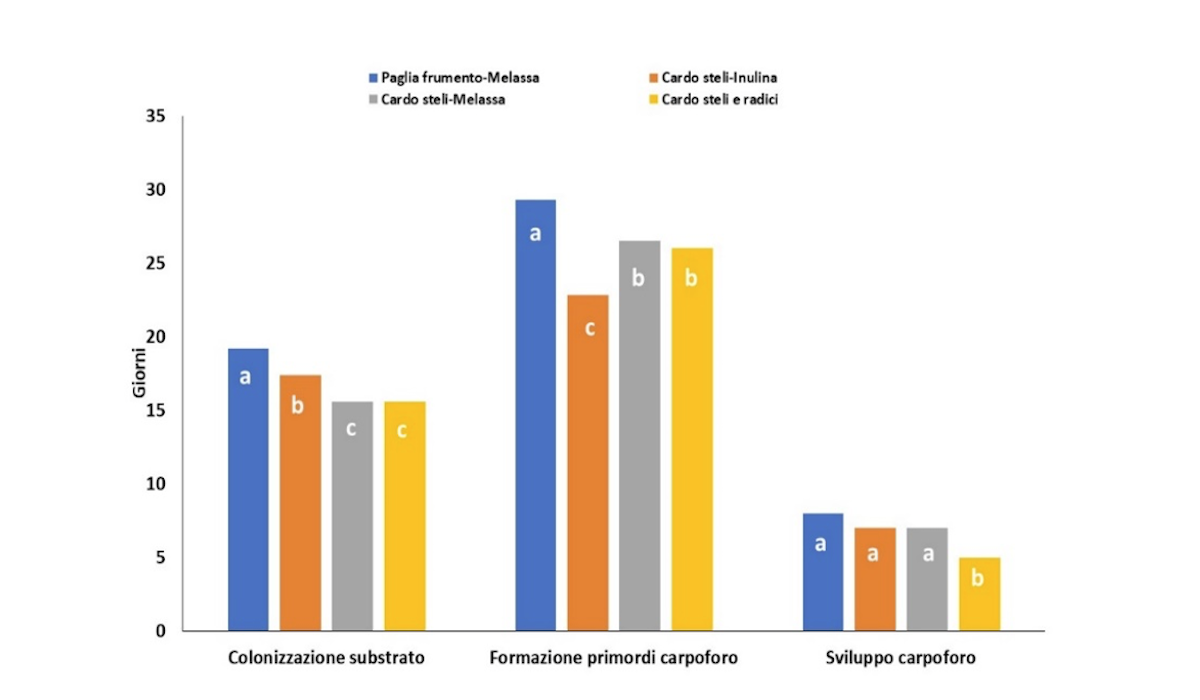
Figura 3.A - Influenza sui parametri produttivi di Pleurotus in diversi substrati a base di cardo. I valori indicati nel grafico sono la media dei valori di tre saggi con tre repliche tecniche. Le lettere indicano le differenze significative fra i trattamenti, valutate mediante analisi della varianza (ANOVA) e test di Tukey con p ≤ 0.05
(Fonte: Crea)

Figura 3.B - Influenza dell'utilizzo delle radici esauste di cardo (a sinistra) sulla velocità di colonizzazione del substrato da parte del micelio di Pleurotus eryngii isolato Pe1CE
(Fonte: Crea)
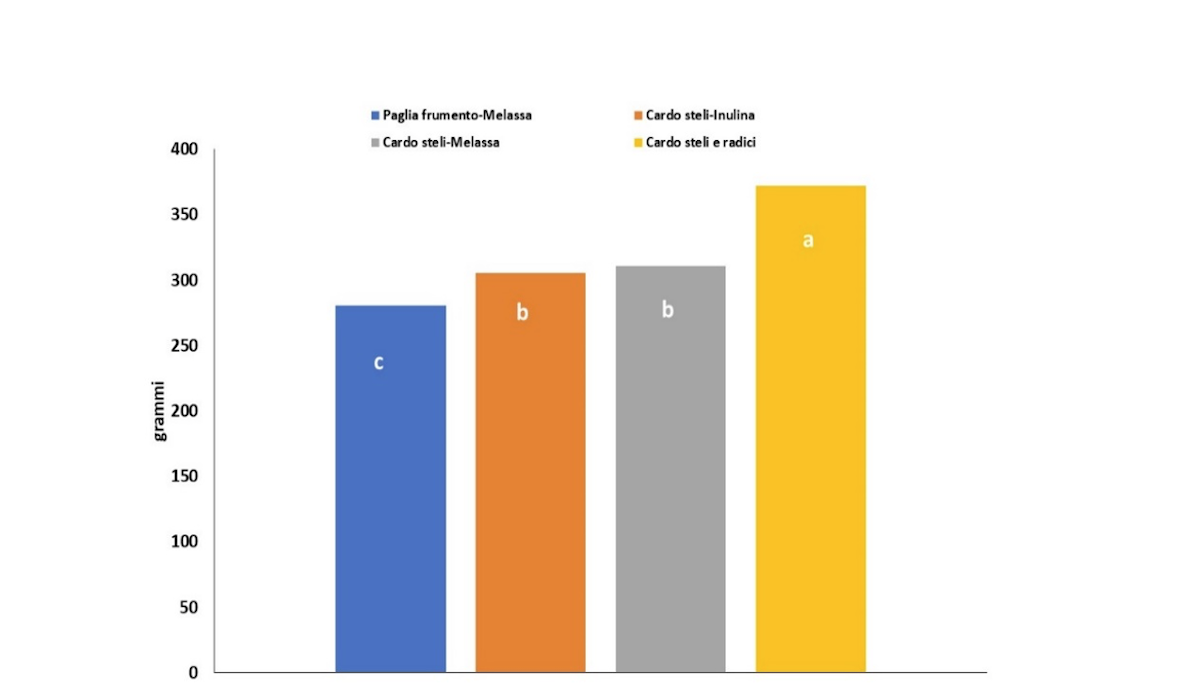
Figura 4.A - Produzione di carpofori di Pleurotus in diversi substrati di coltivazione. I valori indicati nel grafico sono la media dei valori di tre saggi con tre repliche tecniche. Le lettere indicano le differenze significative fra i trattamenti, valutate mediante analisi della varianza (Anova) e test di Tukey con p ≤ 0.05
(Fonte: Crea)

Figura 4.B - Produzione di carpofori di Pleurotus eryngii isolato Pe1CE su substrato paglia di frumento-Melassa (sinistra) e su steli e radici esauste di cardo
(Fonte: Crea)
Contenuto di eteroglicani nei carpofori e nel micelio di P. eryngii
La figura 5 riporta la correlazione positiva del contenuto in eteroglicani nel micelio e nei carpofori di P. eryngii ottenuti in presenza di inulina esogena o presente nella biomassa di cardo. I risultati mostrano che, quando si aggiunge inulina esogena al substrato, nel micelio di P.eryngii vi è maggiore quantità di eteroglicani (1706,3 µg g-1) rispetto al substrato contenente saccarosio (739,7 µg g-1). Inoltre, il contenuto di eteroglicani è anche maggiore rispetto a G. lucidum (468,2 µg g-1).
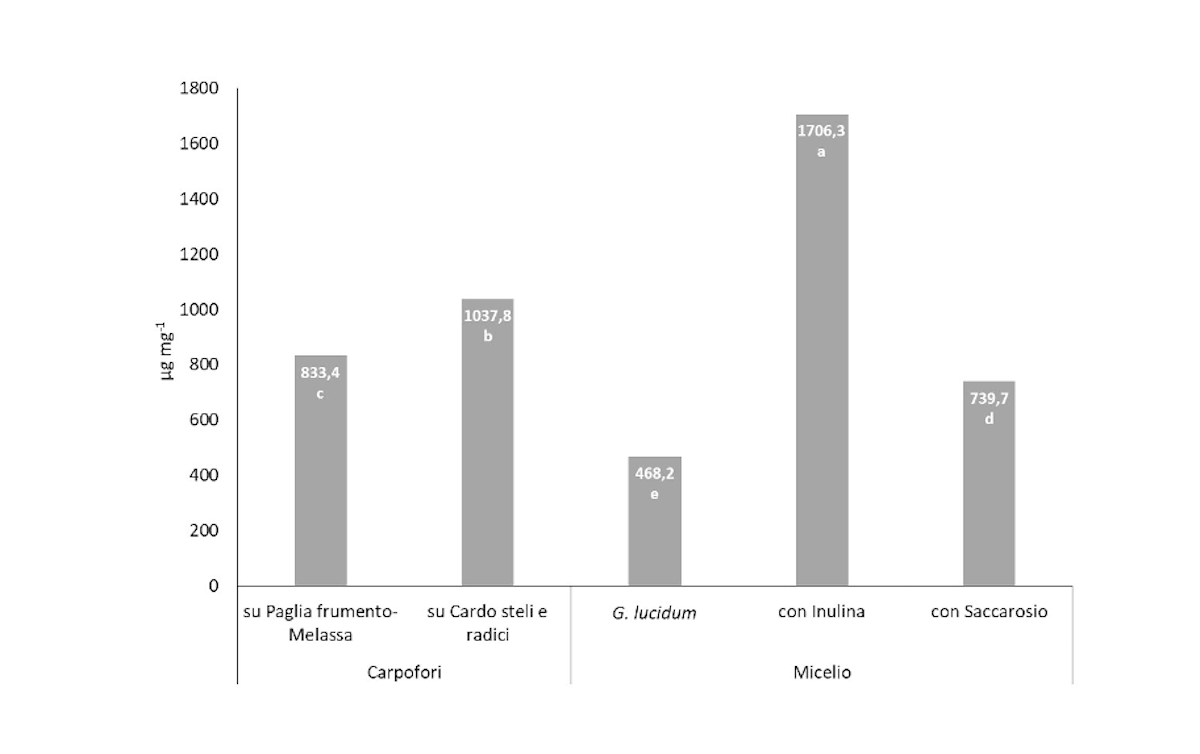
Figura 5 - Contenuto di eteroglicani nei carpofori cresciuti su Pfm (Paglia frumento-Melassa) e CSR (Cardo Steli-Radici) di P. eryngii e G. lucidum. I dati rappresentano i valori medi di tre esperimenti con tre repliche tecniche. Le medie con la stessa lettera non sono significativamente diverse secondo il test di Tukey per p ≤ 0,05
(Fonte: Crea)
I carpofori di P. eryngii coltivati su Csr hanno mostrato un contenuto di eteroglicani più elevato (1037,8 µg g-1) rispetto a quelli ottenuti su Pfm (833,4 µg g-1), ciò potrebbe essere imputato all'assenza della biomassa di cardo.
Valutazione di diversi composti bioattivi e dell'attività antiossidante e degli enzimi coinvolti nella reazione di imbrunimento e nel danno di membrana
Il contenuto dei composti bioattivi, l'attività antiossidante e gli enzimi coinvolti nell'imbrunimento e nel danno alle membrane sono riportati nella Figura 6.
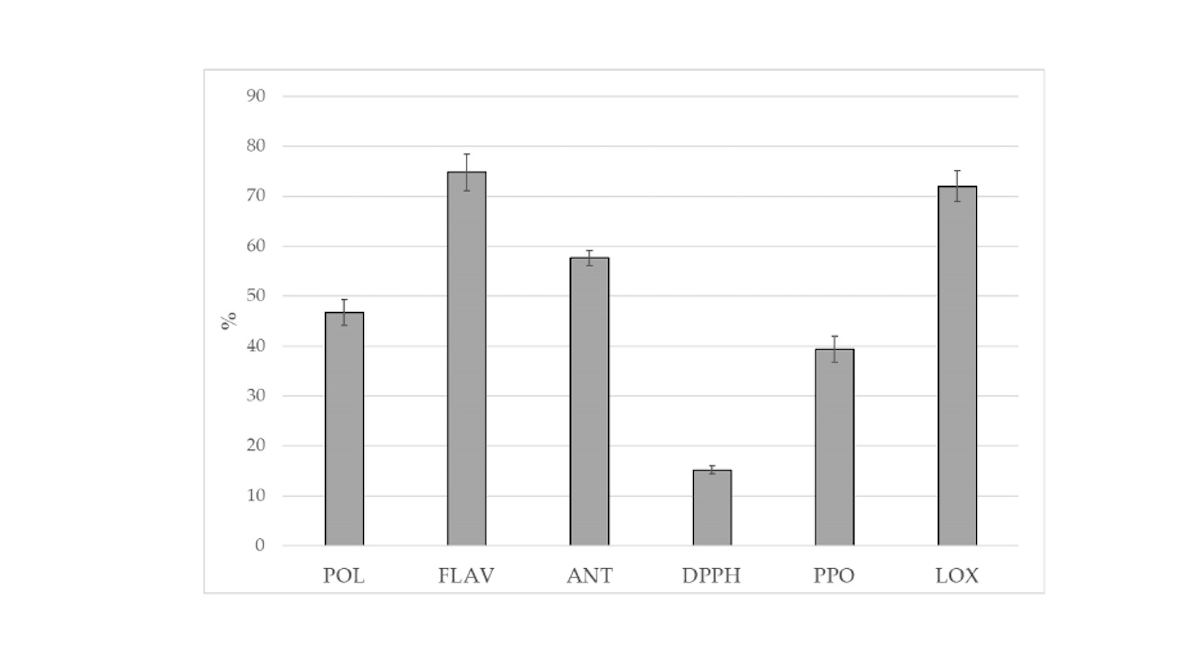
Figure 6 - Incremento percentuale del contenuto di polifenoli (Pol), flavonoidi (Flav), antocianine (Ant), attività antiossidante (Dpph), attività della polifenolossidasi (Ppo) e della lipossigenasi (Lox) in P. eringy ottenuto dal substrato di crescita Pfm rispetto a Csr. I dati rappresentano la media di tre esperimenti con tre replicati tecnici
(Fonte: Crea)
I carpofori ottenuti dal substrato Csr mostrano un incremento del 47% nel contenuto di polifenoli del 75% in flavonoidi rispetto al campione Pfm (Figura 3). Analogamente anche il contenuto di antocianine (57%) risulta incrementato e conseguentemente l'attività antiossidante 815%. Le attività della Ppo e Lox correlate all'imbrunimento enzimatico e al danno ossidativo nei due campioni di funghi mostrano un incremento pari al 39 e 72% (Figura 3).
Risultati del compostaggio
Nella figura 7 è rappresentato l'andamento della temperatura massima e di quella media misurata all'interno del cumulo durante l'intero processo. Nei primi 15 giorni è stato raggiunto e mantenuto una temperatura mediamente superiore ai 50 gradi con diversi picchi tra 55 e 60 gradi. Questa è la fase termofila del processo in cui avviene anche la igienizzazione della massa proprio grazie alle alte temperature per effetto dell'attività microbica a carico delle frazioni organiche più labili.
I tre picchi termici che si osservano durante la fase di maturazione del cumulo sono conseguenti ai rivoltamenti manuali e indicano un rialzo dell'attività microbica dovuto al rimescolamento dei materiali. E' evidente, comunque, un andamento calante della temperatura che tende ad avvicinarsi sempre più a quella ambientale.
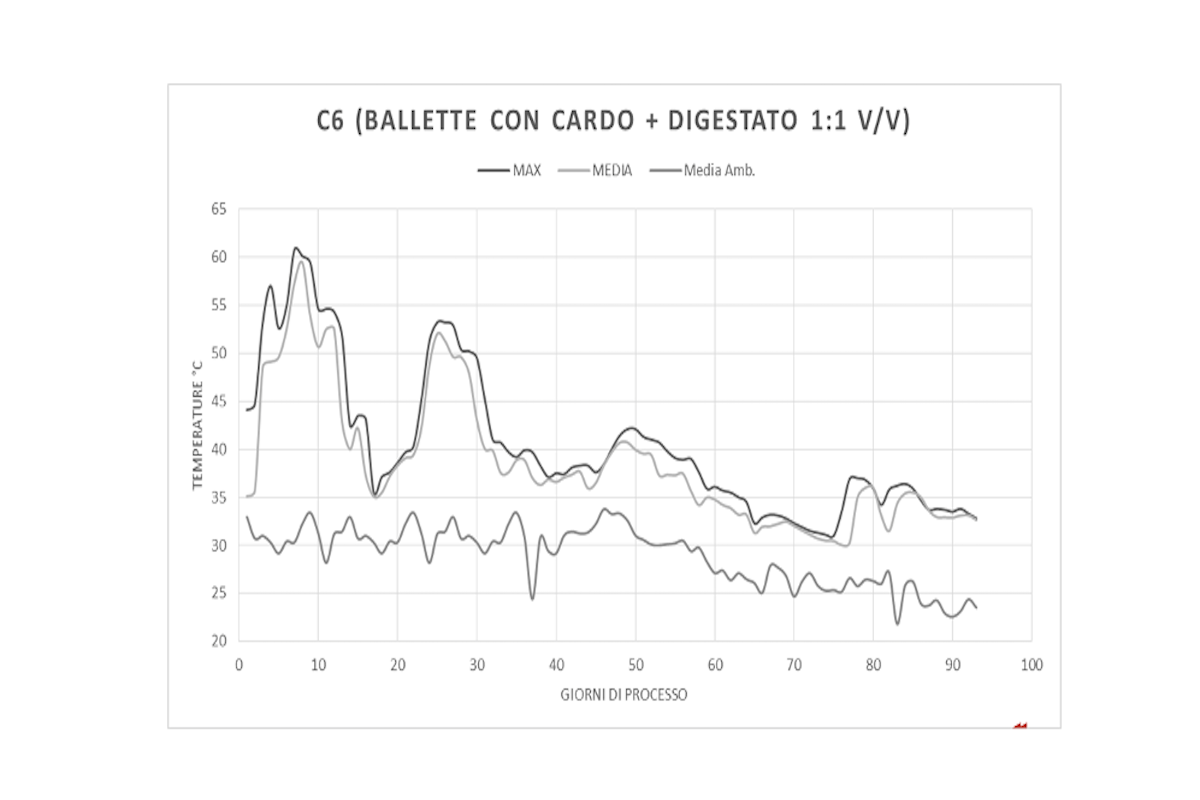
Figura 7 - Andamento della temperatura massima e media a confronto con quella ambientale durante il compostaggio del cumulo con balle esauste da funghicoltura e digestato
(Fonte: Crea)
L'esame dei parametri chimici misurati nei campioni di compost in tre momenti del processo di compostaggio (Tabella 2), ne conferma il buon andamento. Infatti,la concentrazione di C organico diminuisce in conseguenza del consumo microbico e si attesta al 18,7% sul secco; l'N totale invece si concentra venendo riutilizzato dalle popolazioni microbiche che si succedono nelle diverse fasi. Il rapporto C/N, come atteso, si abbassa progressivamente individuando un materiale di pronto effetto una volta immesso nel suolo come ammendante. Infine, pH e conducibilità elettrica restano abbastanza stabili e alti (in particolare la conducibilità) dipendendo direttamente dalle caratteristiche delle matrici di partenza.
Questo processo di compostaggio sperimentale ha dunque dimostrato come sia possibile chiudere, secondo una logica di economia circolare, con la produzione di un fertilizzante organico, una serie di processi produttivi tra loro collegati.
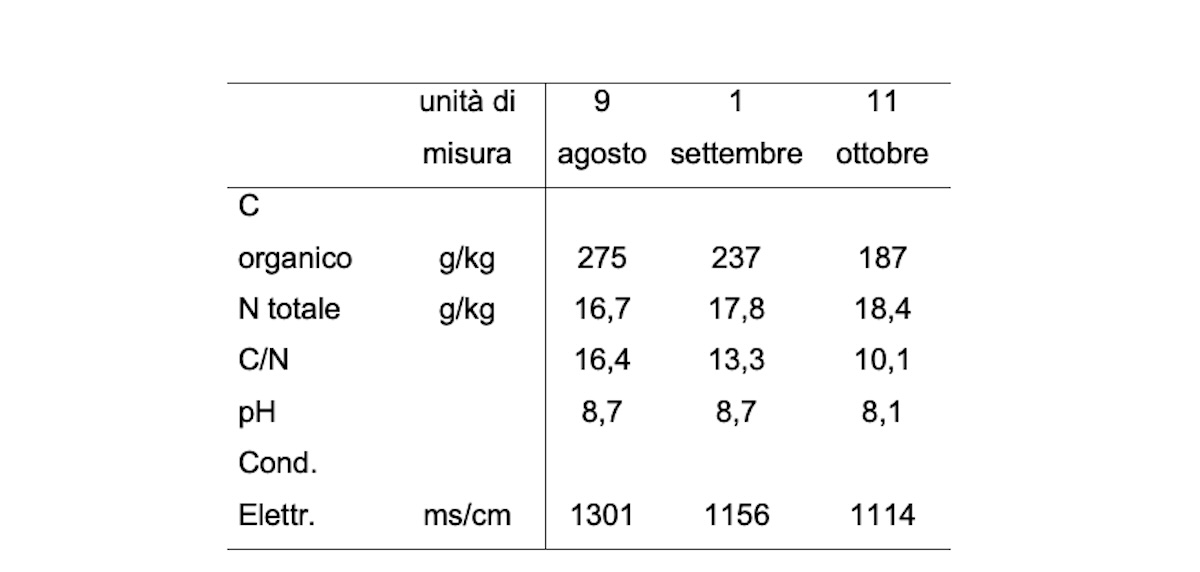
Tabella 2 - Parametri chimici misurati nei campioni di compost in tre momenti del processo di compostaggio
(Fonte: Crea)
Discussione
L'utilizzo della biomassa residua del cardo come substrato per la coltivazione di funghi eduli costituisce un'applicazione innovativa nella filiera di coltivazione di questa specie molto versatile, ben adattata al clima mediterraneo e dalle alte rese potenziali. Si può realizzare così la conversione di un residuo colturale in un prodotto con un valore aggiunto come fonte alimentare alternativa per l'uomo sia nutrizionale che funzionale in quanto naturalmente ricca di molecole e con proprietà benefiche e protettive per l'organismo, se consumata regolarmente e in quantità efficaci nell'ambito di una dieta variata.
Nella sperimentazione condotta si è valutato l'utilizzo dei residui colturali di cardo quale substrato alternativo per la crescita del "cardoncello" la cui richiesta sul mercato, nazionale ed europeo è in crescita e che tra i Pleurotus presenta le migliori qualità nutrizionali.
In particolare sono stati studiati l'affinità, la velocità di crescita e alcune proprietà nutrizionali di cardoncello coltivato utilizzando la biomassa di cardo (steli e radici ricche di inulina esauste o meno) come substrato di crescita.
In primo luogo è stata effettuata la selezione degli isolati di P. eryngii sulla base della loro capacità di crescere su substrati contenenti fusto e radici di cardo, ricchi di inulina, rispetto alla crescita sul comune substrato contenente paglia e melassa (Battaglia et al., 2023). È noto che l'uso di inulina e/o saccarosio come fonti di carbonio consente una crescita più rapida dei funghi. I risultati hanno dimostrato che l'inulina non solo è una fonte di carbonio adatta a sostituire l'uso della melassa per la crescita di P. eryngii, ma anche che, in alte concentrazioni, produce rese maggiori di micelio fungino sia in termini di quantità che di qualità.
Infatti, utilizzando inulina come fonte di carbonio è stata evidenziata una maggiore produzione di eteroglicani nel micelio e nel carpoforo.
Recentemente, lo sviluppo di alimenti o farmaci funzionali contenenti polisaccaridi fungini sta riscuotendo sempre più interesse (Aida et al., 2009). Ad esempio il G. lucidum, fungo con potenti proprietà farmacologiche, è ampiamente utilizzato su scala industriale per la produzione di integratori alimentari contenenti beta-glucano e può essere coltivato in un bioreattore per produrre micelio ed estrarre molecole funzionali quali eteroglicani ed eteroglicani idrosolubili (Carrieri et al., 2017).
Nella presente sperimentazione, il contenuto di eteroglicani ottenuto dal micelio di G. lucidum, utilizzato come controllo, è risultato notevolmente inferiore rispetto a quello di P. eryngii. indicando che il micelio di P. eryngii ottenuto con substrati ad alta concentrazione di inulina, è un eccellente produttore di eteroglicani, sia per resa che per velocità di produzione.
P. eryngii è stata segnalata come la specie di Pleurotus con il più alto contenuto di glucani e pertanto potrebbe essere utilizzata come fonte nutritiva ad alta apporto di glucani (Avni et al., 2017).
Negli ultimi anni, la coltivazione di cardo con svariati scopi energetici ed industriali si è andata espandendo negli ambienti dell'Italia meridionale con conseguente incremento della disponibilità dei suoi residui colturali; pertanto, nel lavoro si è utilizzata questa biomassa per coltivare P. eryngii e ottenere corpi fruttiferi anche come alimento funzionale. I residui colturali del cardo sono anche una fonte naturale di inulina nelle radici (Raccuia et al., 2004) da qui la scelta, in linea con i principi dell'economia circolare, di utilizzare i fusti e le radici rispettivamente come fonte di lignina-cellulosa e inulina per la coltivazione del cardoncello, anche come alimento funzionale.
Diverse tipologie di biomasse di scarto di origine vegetale come caffè, segatura, bagassa di canna da zucchero, paglia di riso e paglia di banana (Tarko et al., 2018) sono state sperimentate per la coltivazione di Pleurotus spp., ma questa è la prima segnalazione sulla possibilità di produrre il cardoncello su residui di cardo. In questo lavoro il miglior substrato di crescita con steli e radici di cardo ha permesso un incremento del 25% del peso medio dei corpi fruttiferi rispetto a quello tradizionale (Pfm), e del 15% rispetto a quanto riportato per il caffè, evidenziato quale migliore tra i substrati alternativi. I corpi fruttiferi ottenuti su residui di cardo (Csr) hanno anche mostrato elevate proprietà organolettiche e nutrizionali.
In particolare, è stata evidenziata un'elevata quantità di composti bioattivi ed elevata attività antiossidante oltre che una maggiore quantità di eteroglicano rispetto al Pfm. Il contenuto in polifenoli totali (Tpc) del cardoncello è risultato maggiore del 50% se coltivato su residui di cardo (Csr) rispetto al comune substrato contenente paglia e melassa (Pfm); i valori determinati sono superiori anche a quelli riportati per i carpofori ottenuti con altri substrati alternativi come biomassa residua di cotone o segatura (Sardar et al., 2017). I valori elevati di polifenoli potrebbero essere dovuti al notevole apporto di nutrienti dal substrato, che migliora il sistema antiossidante non enzimatico coinvolto nella rimozione dei radicali liberi e nell'attivazione degli enzimi antiossidanti. Anche per il contenuto in flavonoidi ed antociani è stato osservato un incremento nel carpoforo coltivato sul substrato Csr rispetto a quello tradizionale (Pfm).
Il contenuto di composti fenolici è un importante indicatore della capacità antiossidante con una stretta correlazione tra queste due variabili. In letteratura è riportato che P. eryngii ha mostrato una maggiore attività antiossidante in termini di capacità di rimozione dei radicali liberi, potenzialmente dannosi per le strutture cellulari, rispetto ad altri funghi dello stesso genere (Mishra et al., 2013).
Per quanto riguarda le molecole funzionali, va evidenziato che i carpofori ottenuti dal substrato Csr presentano un incremento del contenuto di eteroglicani del 20,3% rispetto al substrato di crescita tradizionale Pfm.
I parametri nutrizionali valutati indicano che la scelta del substrato di crescita è adatta per ottenere corpi fruttiferi con un elevato valore nutraceutico e funzionale. La novità di questo studio è rappresentata dall'utilizzo di un residuo colturale con una composizione biochimica della biomassa che copre tutti i fabbisogni nutrizionali di P. eryngii e con radici ricche di inulina utilizzabili come fonte di carbonio.
L'insieme dei risultati ottenuti indica che i residui di biomassa del cardo costituiscono una scelta eccellente come substrato per coltivazione del fungo commestibile P. eryngii.
Altro aspetto riguarda la produzione di inulina dalle radici e l'utilizzo della stessa. Dalle radici si può dapprima estrarre l'inulina da commercializzare come prodotto prebiotico o da indirizzare ai bioreattori come fonte di carbonio per la produzione rapida di micelio di P.eryngii da cui estrarre glucani; le radici esauste dopo estrazione (contenenti ancora buone quantità di inulina) saranno indirizzate insieme con gli steli alla produzione di carpofori.
A conclusione del ciclo produttivo del cardoncello i panetti di substrato esausti, ancora ricchi di sostanze organiche, tra cui micelio fungino, sono stati sottoposti a compostaggio per la produzione di ammendante organico di buona qualità.
Il modello produttivo presentato, generato a valle della coltivazione del cardo, esemplifica il concetto di un circuito produttivo connotato da un virtuoso flusso delle risorse. Lo sviluppo di colture non food a basso input, come il cardo, può produrre ricadute ambientali, sociali ed economiche positive nelle aree interne marginali del Mezzogiorno d'Italia alimentando circuiti produttivi per il comparto agricolo e agroindustriale in linea con i principi di economia circolare.
Finanziamento
Questa ricerca è stata finanziata da "Colture autoctone mediterranee e loro valorizzazione con tecnologie avanzate di chimica verde" - acronimo COMETA. Ministero dell'Istruzione, dell'Università e della Ricerca ARS01_00606.
A cura di Battaglia Valerio, Sorrentino Roberto, Cermola Michele, Morra Luigi, del Piano Luisa e Lahoz Ernesto, Crea - Centro di ricerca per la cerealicoltura e le colture industriali
Petriccione Milena, Crea - Centro di ricerca olivicoltura, frutticoltura e agrumicoltura
Bibliografia
Aida, F.M.N.A.; Shuhaimi, M.; Yazid, M.; Maaruf, A.G. Mushroom as a potential source of prebiotics: A review. Trends Food Sci. Technol. 2009, 20, 567-575.
Avni, S.; Ezove, N.; Hanani, H.; Yadid, I.; Karpovsky, M.; Hayby, H.; Gover, O.; Hadar, Y.; Schwartz, B.; Danay, O. Olive Mill Waste Enhances α-Glucan Content in the Edible Mushroom Pleurotus eryngii. Int. J. Mol. Sci. 2017, 18, 1564.
Battaglia Valerio, Sorrentino Roberto, Verrilli Giulia, del Piano Luisa, Sorrentino Maria Cristina, Petriccione Milena, Sicignano Mariarosaria, Magri Anna, Cermola Michele, Cerrato Domenico and Lahoz Ernesto. Potential Use of Cardunculus Biomass on Pleurotus eryngii Production: Heteroglycans Content and Nutritional Properties (Preliminary Results). Foods 2023, 12, 58.
Carrieri, R.; Manco, R.; Sapio, D.; Iannaccone, M.; Fulgione, A.; Papaianni, M.; de Falco, B.; Grauso, L.; Tarantino, P.; Ianniello, F. Structural data and immunomodulatory properties of a water-soluble heteroglycan extracted from the mycelium of an Italian isolate of Ganoderma lucidum. Nat. Prod. Res. 2017, 31, 2119-2125.
Mishra, K.K.; Pal, R.S.; Arunkumar, R.; Chandrashekara, C.; Jain, S.K.; Bhatt, J.C. Antioxidant properties of different edible mushroom species and increased bioconversion efficiency of Pleurotus eryngii using locally available casing materials. Food Chem. 2013, 138, 1557-1563.
Nitschke, J.; Modick, H.; Busch, E.; von Rekowski, R.W.; Altenbach, H.-J.; Mölleken, H. A new colorimetric method to quantify-1,3-1,6-glucans in comparison with total -1,3-glucans in edible mushrooms. Food Chem. 2011, 127, 791-796.
Raccuia, S.A.; Melilli, M. Cynara cardunculus L., a potential source of inulin in the Mediterranean environment: Screening of genetic variability. Aust. J. Agric. Res. 2004, 55, 693-698.
Sardar, H.; Ali, M.A.; Anjum, M.A.; Nawaz, F.; Hussain, S.; Naz, S.; Karimi, S.M. Agro-industrial residues influence mineral elements accumulation and nutritional composition of king oyster mushroom (Pleurotus eryngii). Sci. Hortic. 2017, 225, 327-334.
Tarko, D.B.; Sirna, A.M. Substrate optimization for cultivation of Pleurotus ostreatus on lignocellulosic wastes (coffee, sawdust, and sugarcane bagasse) in Mizan-Tepi University, Tepi Campus, Tepi Town. J. Appl. Biol. Biotechnol. 2018, 6, 14-20.
© AgroNotizie - riproduzione riservata
Fonte: Crea - Consiglio per la ricerca in agricoltura e l’analisi dell’economia agraria




















